| [Accueil] |
 |
|||
|
III. Le Système nerveux périphérique
Les nerfs périphériques assurent la transmission de l'information :
- depuis des structures réceptrices périphériques vers le système nerveux central : ce sont les neurones périphériques sensitifs ou sensoriels.
- depuis le système nerveux central vers les organes cibles : ce sont les neurones moteurs.
Les nerfs rachidiens ont une répartition selon les "SOMITES PRIMITIFS" du corps humain : chacun distribue ses neurones à une région précise du corps humain.
Les neurones sensitifs, moteurs et végétatifs sont mélangés dans le nerf rachidien. Ils se séparent au niveau des racines des nerfs, les éléments sensitifs devenant postérieurs et les moteurs antérieurs. Il y a donc en ARRIERE les racines SENSITIVES et en AVANT les racines motrices. Les corps cellulaires des neurones sensitifs sont tous regroupés au niveau du "ganglion rachidien" situé sur la racine postérieure du nerf.
1° Les informations sensitivo-sensorielles véhiculées vers le système nerveux sont d'origine variée :
- fibres libres du tissu sous-cutané ou des organes : douleur
- organes périphériques spécialisés dans la capture des informations venues de l'extérieur (extéroceptives) (disques de Merkel = toucher léger / corpuscules de Meissner = toucher appuyé / corpuscules de Ruffini = chaleur / corpuscules de Krausse = froid / corpuscules de Golgi = pressions)
- organes des informations de notre propre corps : informations proprioceptives : tendons, muscles (fuseaux neuro-musculaires), articulations. informations intéroceptives (viscérales).
2° Les efférences motrices sont aussi de destination diverses : muscles squelettiques, muscles lisses, vasomotricité, muscles des poils, système végétatif.
IV. Moelle segmentaire. Réflexe myotatique & polysynaptique
La moelle est constituée de substance blanche [grec Leuko] (connexions neuronales) et de substance grise [grec Polio] (corps cellulaires). La substance grise, en forme de papillon, est située au milieu, entourée de substance blanche. Au centre de la moelle est situé un petit canal (canal épendymaire) rempli de liquide céphalo-rachidien, communiquant en haut avec les ventricules cérébraux, mais fermé en bas.
Au niveau de la moelle épinière, chaque niveau de racines de nerf rachidien correspond à une zone fonctionnelle autonome. La Moelle est en quelque sorte un "empilement" de zones successives comme les rondelles d'un saucisson (structure dite "métamérique").
Dans la moelle comme dans l'encéphale, les structures sensitivo-sensorielles sont POSTERIEURES, les structures végétatives sont MEDIANES, les structures motrices sont ANTERIEURES.
Les informations sensitives, à leur arrivée dans la moelle, se séparent en 2 grands contingents :
- informations à valeur thermique et douloureuse.
- informations concernant le tact et la proprioception (sensibilité interne à notre corps).
Les informations thermo-algiques font relais dès leur entrée dans la moelle, au niveau de la corne postérieure de la substance grise. Ce relais des voies de la douleur permet un contrôle des informations et leur diffusion.
Entre autres, cette diffusion donne 2 types de connexions :
* Une première diffusion vers les cornes antérieures est à l'origine du réflexe polysynaptique de retrait : grâce à l'existence de ce réflexe, les informations à valeur d'alerte aboutissement rapidement à un évitement (la main posée sur le feu est retirée bien avant que la conscience dise "ouille")
* Une autre collatérale va donner naissance à un "faisceau" d'informations groupées qui va croiser la ligne médiane puis se diriger vers le haut ; c'est le FAISCEAU SPINO-THALAMIQUE, véhiculant ces informations désagréables vers les structures sus-jacentes. La fonction de ce faisceau est aussi une fonction d'alerte, mais destinée aux structures du cerveau.
Les informations proprioceptives donnent elles aussi à l'entrée dans la moelle plusieurs embranchements ("collatérales").
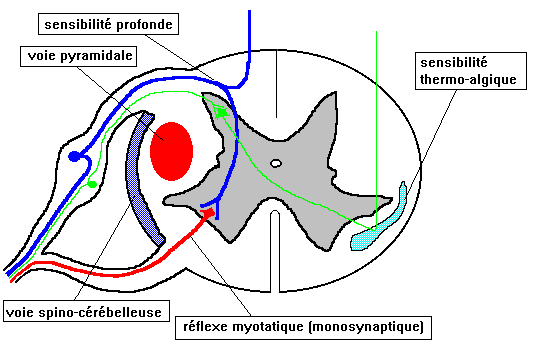
Les informations proprioceptives émanant d'un organe musculaire appelé "fuseau neuro-musculaire" donnent une collatérale qui se dirige vers l'avant et rejoint directement le MOTONEURONE MOTEUR de la corne antérieure de la moelle. Cette "BOUCLE" est essentielle dans le fonctionnement de nos muscles striés, TOUTES LES COMMANDES venues de plus haut ne faisant que moduler l'état de repos de ce système : c'est le réflexe myotatique d'étirement (fonction antigravitaire - combat la gravité terrestre - permet de marcher et de tenir debout). En clinique, ce qu'on appelle "réflexes tendineux" et que l'on étudie avec le marteau à réflexes n'est que l'étude de l'état de cette "boucle" neuronale : si la boucle est "hyperexcitable" en raison d'ordres anormaux venus de plus haut, le réflexe sera trop vif (cas des hémiplégies). Si au contraire il y a interruption de la boucle, le réflexe est aboli (cas des polynévrites).
Une autre collatérale du neurone proprioceptif va se diriger SANS RELAI vers le cordon postérieur de la moelle : le groupement de ces neurones constitue une voie amenant vers le cerveau les informations, appelée "faisceaux de Goll et Burdach". Ce faisceau reste du même côté et ne croise dont pas la ligne médiane au niveau de la moelle.
fait fondamental TOUTES LES ACTIONS DES STRUCTURES SUS-JACENTES NE FONT QUE MODULER CES DEUX BOUCLES REFLEXES.
En résumé :
La moelle dite "segmentaire" est une tranche de système nerveux central branchée sur une tranche de notre corps. Elle constitue le relais des entrées et sorties du système nerveux central.
Ce système "entrées et sortie" fonctionne sur 2 "réflexes" fondamentaux : l'un "myotatique", précis, rapide, destiné à nous permettre des gestes fins en tenant compte de la pesanteur. L'autre, "d'évitement", plus diffus, est un système d'alerte dont la fonction est la conservation de l'intégrité du corps en face d'agressions extérieures.
V. Moelle intersegmentaire
A. VOIES D'ASSOCIATION
Comme cela a été vu au chapitre précédent, les neurones sensitifs en entrant dans la moelle se divisent en nombreuses ramifications assurant la diffusion de l'information.
Il existe des neurones spécialisés pour cette fonction de diffusion, assurant le transfert des informations d'un niveau à l'autre de la moelle : ce sont les cellules cordonnales : leurs prolongements sont plaqués autour de la substance grise, et constituent ce que l'on appelle le faisceau fondamental de la moelle (antéro-latéral) et la zone commissurale (postérieure).
B. CENTRES MEDULLAIRES PLURISEGMENTAIRES
1. CENTRES DE LA CORNE ANTERIEURE
Il ne faut pas voir les structures donnant naissance aux nerfs rachidiens comme strictement localisées à la région d'origine du nerf. Les noyaux des nerfs rachidiens forment des colonnes réparties en hauteur, ces colonnes étant toujours situées immédiatement au dessus de l'émergence de la racine concernée.
- C1 à C8 : colonnes des centres musculaires de la nuque et du cou : muscles de l'épaule C4-C7, bras C5-C8, avant-bras C6-C8, main C7-D1 (cas particulier du "centre spinal médullaire" = trapèze et sterno-cleido-mastoïdien : émergence très originale devant les racines postérieures noyau C1-C6 antéro-externe).
- C5 à D12 : muscles du thorax
- D5 à coccyx : parois abdominales et pelviennes.
- Muscles de la loge antérieure de cuisse L2-L4. Face postérieure de cuisse L4-S3.
2. CENTRES DE LA CORNE POSTERIEURE
Agencement probable en colonnes (colonne de Clarke = premier relais de la voie proprioceptive inconsciente). Beaucoup moins clair que pour les centres moteurs de la corne antérieure.
VI. Moelle : lieu de transit des informations
A. Faisceaux ascendants
1. Voies extéroceptives
- Sensibilité extéroceptive nociceptive : voie de la température et de la douleur faisceau spino-thalamique. Associé au tact protopathique ("grossier"). Le faisceau spino thalamique monte vers l'encéphale dans le cordon antéro-latéral de la moelle. Il perd au cours de son trajet de nombreuses collatérales. Il y a donc concentration des informations douloureuses, expliquant que la douleur est une perception diffuse, difficile à situer sur le corps.
- Sensibilité extéroceptive discriminative (tact épicritique "précis") Cordon postérieur de la moelle. Ce cordon précis constitué de neurones à vitesse de conduction rapide est mis en jeu dans toutes les adaptations fines des positions des membres face à un besoin. Il véhicule des informations concernant la position des membres dans l'espace, sans ces informations le cerveau sera incapable de donner des ordres précis pour un mouvement, surtout si il n'y a plus de contrôle de la vision. Cette sensibilité s'étudie donc en clinique les yeux fermés, le malade se montrant incapable, dans ces conditions, de savoir où est situé son membre.
2. Voies proprioceptives
Les voies de la sensibilité profonde sont à la fois conscientes et inconscientes.
- Inconscientes : faisceaux spino-cérébelleux direct et croisé, situés latéralement dans la moelle, portant les informations vers le cervelet.
- Conscientes : fibres bulbopètes (c-à-d allant vers le bulbe) des cordons postérieurs, nous l'avons déjà vu.
3. Voies intéroceptives
Elles apportent vers le système nerveux central des informations venant des viscères.
La "Substance grise végétative périépendymaire" ayant une "fonction conductrice propre", il n'y a pas à proprement parler de voie sensitive pour le système nerveux végétatif.
Cependant, les afférences d'origine profonde véhiculées par les nerfs proprioceptifs (afférences "intéroceptives") ont aussi des impressions nociceptives : "douleurs viscérales", perçues de façon très diffuse, mal localisée, parfois même projetées en raison de groupements neuronaux ; c'est le cas des douleurs d'origine vésiculaire, volontiers localisées à l'épaule, ou des douleurs pancréatiques qui sont perçues dans le dos, des douleurs de l'infarctus myocardique senties dans le bras gauche,...
B. Faisceaux descendants
1. Voie pyramidale
La voie dite "pyramidale" est aussi appelée "voie motrice principale". Cette voie est constituée des neurones traversant une partie anatomique du bulbe rachidien appelée "pyramide bulbaire", d'où son nom. Par définition, toutes les voies motrices qui n'empruntent pas ce faisceau "pyramidal" sont appelées "extra-pyramidales" (ces dernières sont numériquement beaucoup plus importantes que la voie "pyramidale").
Le faisceau pyramidal est aussi appelé "faisceau cortico-spinal" ou "pyramidal croisé" (il existe aussi un faisceau "pyramidal direct" qui est tout à fait négligeable dans l'espèce humaine). Au niveau de la moelle, il est situé dans le cordon latéral de la substance blanche, juste en dehors de la corne antérieure.
Son atteinte en pathologie provoque une paralysie d'origine centrale particulière appelée "syndrome pyramidal". C'est ce type de paralysie qui constitue l'hémiplégie.
2. Voies extrapyramidales
Faisceaux descendants formés à tous les étages de l'encéphale : réticulo-spinal, vestibulo-spinal.
Ces voies apportent vers la corne antérieure de la moelle des informations motrices venues du cerveau, modulant en permanence l'état de la boucle du réflexe myotatique. Leurs projections au niveau de la corne antérieure de la moelle se font sur le "motoneurone gamma", petit neurone chargé de régler le "niveau de repos" de la tension du fuseau neuro-musculaire du muscle. Pour comparer ce système à un piano, le système extra-pyramidal règle l'état de tension des cordes et le système pyramidal est le clavier qui permet de jouer : les deux sont donc complémentaires, le système pyramidal jouera "faux" si le système extra-pyramidal ne fonctionne pas normalement.
3. Voies végétatives
Ces voies végétatives sont très importantes pour la vie inconsciente. Elles régulent la sueur, le rythme cardiaque,... C'est à cause d'elles que l'angoisse d'un examen déclenche des sueurs, une accélération du coeur et une diarrhée.
Les faisceaux descendants sont formés dans le tronc cérébral. Ils portent les informations efférentes de l'hypothalamus et des centres végétatifs du tronc cérébral.
VII. Le système nerveux autonome
Les centres végétatifs sont de répartition centrale. (aires végétatives réparties depuis le diencéphale). Neurones groupés en 'bancs de poissons'. Ces centres sont autour des structures médianes dans l'encéphale, et à la partie médiane de la substance grise de la moelle.
Les centres parasympathiques sont situés en haut (encéphale) et en bas (fin de la moelle). Les centres sympathiques (ou "orthosympathique") sont répartis sur toute la hauteur de la moelle épinière.
Le terme de nerfs "sympathiques" vient du fait constaté par les anciens anatomistes que ces nerfs avaient des "communications très fréquentes" avec la plupart des autres nerfs : ils "entraient en sympathie" avec les autres. La chaîne ganglionnaire paravertébrale était appelée "grand nerf sympathique".
Nerfs parasympathiques : le nerf moteur oculaire commun (III) véhicule la motricité permettant la contraction de la pupille. Les nerfs V (trijumeau), VII (facial), IX (glosso-pharyngien) transmettent les ordres aux glandes salivaires. Le nerf pneumogastrique (X) assure une vaste innervation destinée au coeur, aux poumons, au tube digestif). Les centres parasympathiques pelviens sont destinés à l'appareil génito-urinaire. Les voies parasympathiques comprennent un relais (ganglion parasympathique) situé non loin de l'organe cible.
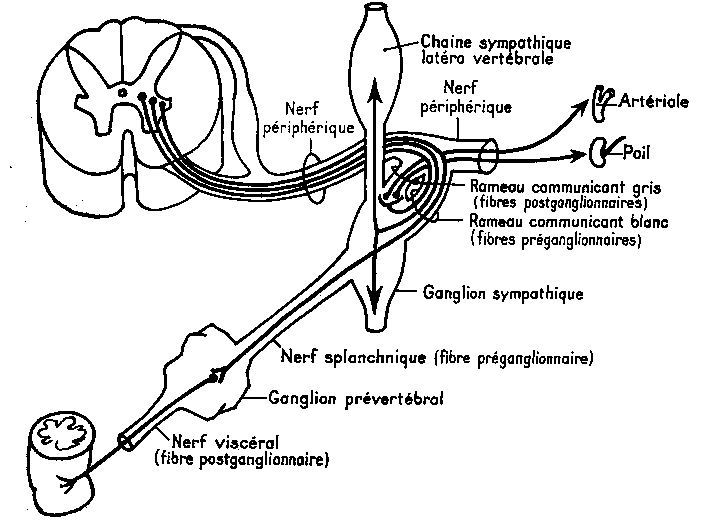
Nerfs sympathiques : Centre cilio-spinal C8-D1, Centre broncho-pulmonaire D3-D5, centres splanchniques abdominaux D6-D9/D10/D12, centres splanchniques pelviens D12-L2 ; relais au niveau du sympathique para-vertébral (chaîne ganglionnaire sympathique proche du système nerveux central).
Le neurone effecteur Viscéral est situé hors du névraxe dans le ganglion. Le trajet effecteur est : centre médullaire è Racine motrice è rameau communicant blanc è ganglion
(Sympathique) èrameau communicant gris ènerf périphérique è organe cible
(Parasympathique) è nerf splanchnique è ganglion prévertébral è fibre post ganglionnaire è organe cible.
D'une façon générale les ganglions sympathiques sont situés près du système nerveux central, les ganglions parasympathiques près des organes cibles. |
|||